
Os herbicidas inibidores do fotossistema II (FSII) atuam nos cloroplastos de células e estão envolvidos na inibição da fotossíntese, porém não matam as plantas por falta de energia, e sim por uma sequência de eventos que veremos logo à frente.
Os herbicidas do grupo C1 e C2 podem ser aplicados tanto no solo quanto em pós-emergência. Quando em pós-emergência, são produtos de reduzida translocação e que necessitam adequada cobertura foliar e uso de adjuvantes. Os do grupo C3 são de contato, não translocam na planta e são usados na modalidade de pós-emergência.
Como atuam os herbicidas inibidores FS II
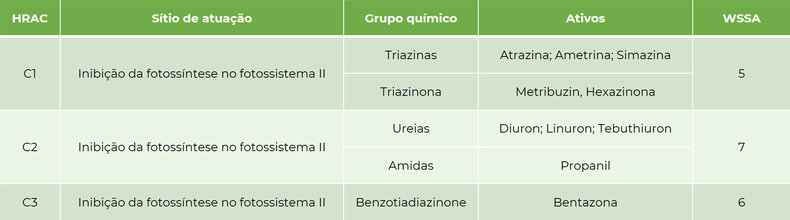
As células fotossintetizantes são aquelas com presença de cloroplastos e consequentemente pigmentos, como as clorofilas. Através da fotossíntese as plantas utilizam a energia da luz para converter CO2 (gás carbônico) e água em energia na forma de glicose e oxigênio. Dentro dos cloroplastos, nas membranas dos tilacoides, encontram-se os fotossistemas, os quais são formados por um sistema de captação (pigmentos) capazes de capturar a energia da luz e transmiti-la para transportadores na forma de elétrons, que serão utilizados para a formação de energia na forma de ATP e NADPH+.
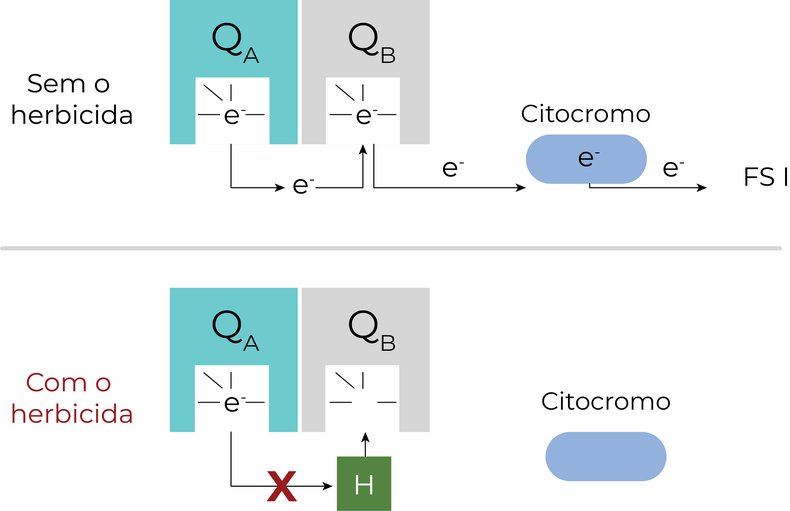
Os elétrons em estado excitado passam por um percurso, entre doadores e receptores. Nesse percurso, no fotossistema II, uma molécula de plastoquinona (QA) ligada a uma proteína (D1) passa elétrons para outra plastoquinona (QB). A QB precisa receber dois elétrons de QA para se tornar protonada, e então transferir elétrons para um citocromo, que por sua vez, transfere elétrons para o fotossistema I.
Os herbicidas inibidores do FSII se ligam ao sítio de ligação de QB, impedindo que essa receba os elétrons de QA, interrompendo o fluxo de elétrons do fotossistema II.
Como se dá a morte das plantas?
Os primeiros trabalhos sugeriam que a morte das plantas se dava por “inanição”, ou seja, falta de alimento (“energia”). No entanto, atualmente se sabe que não é apenas por esse evento que as plantas morrem.
Quando o fluxo de elétrons é interrompido, as moléculas de clorofilas continuam captando energia solar, acumulando energia. Essas clorofilas acabam se tornando altamente excitadas, em um estado chamado de “tripleto”. Esta energia em excesso fica disponível para outros receptores, como o oxigênio, sempre presente nas plantas, por exemplo.
Quando o oxigênio recebe a energia em excesso das clorofilas, são formadas espécies reativas de oxigênio (EROs), conhecidas como radicais livres. O acúmulo desses radicais livres causa sérios danos às células da planta, através principalmente da peroxidação lipídica que danifica as membranas celulares.
A peroxidação de lipídios é um processo que desfaz a organização da membrana celular, causando seu rompimento e consequente extravasamento de seu conteúdo. Após este processo, ocorre a morte celular e com isso surgem os sintomas necróticos nas folhas, característicos da ação deste grupo de herbicidas. Como a peroxidação dos lipídios é autocatalítica, são reações que se espalham nos tecidos, e isso contribui para que este leve a planta a morte.
Efeito da presença de luz na ação desses herbicidas
Essa relação é bem conhecida para herbicidas do grupo das triazinas aplicados via foliar (pós-emergência). Para atrazina, por exemplo, a translocação na planta é bastante limitada, principalmente quando o herbicida é aplicado na presença de luz, pois como a fotossíntese está ativa, os herbicidas iniciam suas reações logo após o contato com as plantas.
Nesse contexto, alguns trabalhos indicam uma melhoria de performance desses herbicidas quando aplicados à noite, já que a noite a fotossíntese não está ativa, podendo estes serem mais eficientemente translocados pelas plantas. Desta forma, no início do dia quando na presença de luz, o herbicida já estará “espalhado” de forma mais uniforme na planta e sua ação será mais efetiva.
Seletividade em milho e sorgo
As plantas de milho e sorgo são tolerantes a triazinas como a atrazina e simazina, pelo processo de detoxificação. Os herbicidas são degradados na planta antes de chegarem aos cloroplastos. Essa degradação ocorre especialmente pelo processo de conjugação com o complexo enzimático glutationa-S-transferase nas folhas.
Outras plantas apresentam capacidade de detoxificação e são também tolerantes a estes herbicidas, como algumas espécies dos gêneros Panicum, Digitaria e Setaria. Outros processos podem contribuir para a tolerância de plantas a esses herbicidas, como por exemplo, a absorção diferencial por raízes e folhas, a translocação diferencial e sorção em sítios inativos nas plantas.



