
Neste material conheça tudo sobre resistência de fungos a fungicidas:
-
as causas da resistência
-
os mecanismos de resistência
-
os tipos de resistência
-
a vulnerabilidade dos diferentes fungicidas
Resistência de fungos a fungicidas
Primeiramente tem-se que diferenciar tolerância de resistência. A tolerância é observada na situação em que uma planta é atacada por um patógeno na mesma intensidade que outras plantas, mas como resultado da infecção, sofre menos danos em termos de produtividade ou qualidade do produto (Robinson, 1969).
A tolerância é controlada por muitos genes e é efetiva contra um número maior de raças do fungo. Normalmente, tem sido obtida por meio da avaliação da produtividade relativa de grãos, comparando parcelas de um mesmo genótipo com e sem proteção por fungicida (Lima, 2012). Já a resistência é uma alteração herdável e estável em um fungo em resposta à aplicação de um fungicida, resultando na redução da sensibilidade ao produto (European, 1988).
Fungos, como todos os organismos vivos, são geneticamente variáveis e podem, através de mutações, tornarem-se resistentes a fungicidas específicos, que atuam em um ou poucos processos metabólicos vitais. As consequências da seleção de indivíduos resistentes a fungicidas podem ser desastrosas, tanto para o usuário, comprometendo sua produção, quanto para o fabricante, que investiu na descoberta e desenvolvimento do produto.
Um gene mutante que causa a produção de um mecanismo de resistência específica preexiste em pequenas quantidades na população. Muito antes de um fungicida ter sido utilizado no campo, esta mutação não ofereceria nenhuma vantagem para o crescimento ou sobrevivência do organismo, e poderia causar uma ligeira desvantagem. Por esta razão, a frequência do gene permaneceria muito baixa, provavelmente desaparecendo e reaparecendo espontaneamente varias vezes (Brent, 1999). As mutações ocorrem por substituição, supressão, adição ou inversão de bases.
Os mecanismos de resistência estão atrelados a modificações no fungo ligadas processos como: desenvolvimento de via metabólica alternativa, degradação metabólica do fungicida, exclusão ou expulsão do fungicida, decréscimo na permeabilidade da membrana, aumento na detoxificação, alteração bioquímica no sítio ativo.
Causas da Resistência
O processo de mutação pode acontecer naturalmente, levando anos para ocorrer ou ser “acelerado” devido, principalmente, a práticas errôneas de manejo químico. As causas da resistência estão ligadas à:
•Cada produto tem um fator inerente de risco;
• Indivíduos resistentes existem em proporção baixa em populações sensíveis;
•O desenvolvimento de resistência depende de pressão de seleção;
•Fatores inerentes ao patógeno contribuem para o surgimento de resistência na população;
•Problemas de tecnologia de aplicação;
•Aplicações tardias;
•Uso de fungicidas isolados;
•Sub ou superdose de um fungicida eficiente.
Mecanismos da Resistência
Os mecanismos de resistência estão ligados à capacidade de seleção de raças resistentes de patógenos, o que pode ser aumentado nos seguintes casos:
(a) aplicação repetida de fungicidas sistêmicos quimicamente relacionados;
(b) aplicação continua de fungicidas em concentrações sub-letais;
(c) utilização de fungicidas em áreas isoladas onde não haja competição das diferentes raças dos organismos.
Decréscimo da permeabilidade: O decréscimo da permeabilidade diz respeito aos fungicidas que não atingem o local de ação devido ao decréscimo na permeabilidade da membrana do patógeno resistente.
Aumento da desintoxicação: O aumento da desintoxicação pode ocorrer por degradação metabólica do fungicida, com concomitante perda de ação do mesmo após a entrada na célula fúngica.
Alteração bioquímica do sítio alvo: A alteração bioquímica do sítio alvo ocorre quando o fungicida alcança o local de ação sem ser metabolizado pelo organismo. Dessa forma, a resistência pode ser explicada baseando-se na falta de afinidade do inibidor no sítio reativo.
Desenvolvimento de via metabólica alternativa: O desenvolvimento de via metabólica alternativa ocorre quando um fungicida bloqueia a reação em um determinado sítio de ação do metabolismo fúngico e o fungo se adapta a esta situação, alterando seu metabolismo de tal forma que o local bloqueado não seja utilizado.
Tipos de Resistência
<
Resistência cruzada: ocorre quando populações de patógenos que desenvolvem resistência a um fungicida tornam-se, automaticamente e simultaneamente, resistentes a outros fungicidas que são afetados pela mesma mutação gênica e pelo mesmo mecanismo de resistência.
Resistência múltipla: é a resistência de linhagens patogênicas que apresentam mecanismos de resistência separados para dois ou mais fungicidas não relacionados.
Resistência qualitativa: é a perda de efetividade do fungicida de modo repentino e marcante pela presença bem definida de populações de patógenos que apresentam suscetibilidade e resistência com respostas que variam amplamente. A resistência qualitativa é monogênica com processo rápido (one step), geralmente irreversível e o fungicida apresenta falha total no controle. A resistência qualitativa também é citada como: resistência ‘estágio único’, resistência ‘discreta’, ‘disruptiva’ ou ‘descontinua’. Uma vez desenvolvida, a tendência da resistência e permanecer estável. Se o fungicida em questão for removido ou usado com menor frequência, as populações de patógenos podem permanecer resistentes por muitos anos. Exemplos: resistência a fungicidas benzimidazois, QoIs e SDHIs.
Resistência quantitativa: tanto o declínio no controle da doença como a diminuição da suscetibilidade das populações do patógeno, demonstradas por teste de monitoramento, se manifestam gradualmente, são parciais e ocorrem em graus variáveis. É uma resistência poligênica com processo gradual (multiple steps), incompleta, passível de reversão e o fungicida apresenta falha parcial de controle. Este tipo de resistência é denominado também por: resistência ‘multi-estágio’, ‘contínuo’, ‘direcional’ ou ‘progressivo’. Esta resistência é rapidamente revertida a uma condição de maior susceptibilidade sob circunstâncias nas quais o fungicida em questão é utilizado de modo menos intensivo e fungicidas alternativos são aplicados contra a mesma doença. Exemplos: resistência a fungicidas DMIs.
Vulnerabilidade dos diferentes fungicidas
Fungicidas Protetores com ação em múltiplos sítios
Agem como um inibidor genérico de enzimas afetando muitos sítios alvo (por isso chamados inibidores ‘multissítios’). Muitos sítios no fungo teriam que sofrer mudanças simultaneamente para impedir a ação do fungicida. Geneticamente, a possibilidade disto acontecer e mínima e, de qualquer modo, um organismo com tantas alterações, dificilmente seria patogênico ou mesmo viável.
Fungicidas Sistêmicos com ação em local específico
Agem primariamente em um unico sítio-alvo e são frequentemente denominados fungicidas de ‘sitio-único’ ou ‘sítio-específico’. Apenas uma mutação em um único gene pode causar uma alteração em um sítio-alvo de modo a se tornar bem menos vulnerável ao fungicida. Presentemente, vários genes maiores foram isolados e caracterizados.
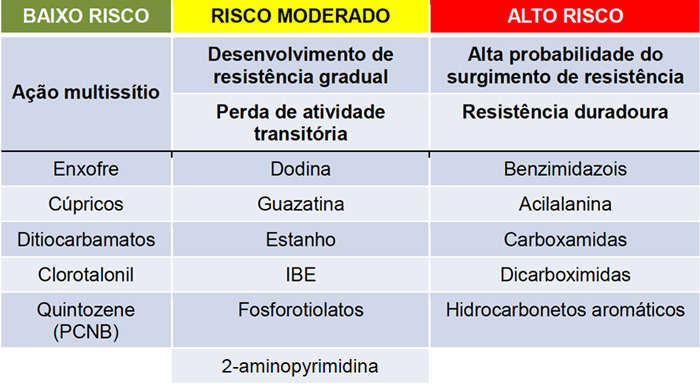
Resistência a fungicidas sítio-específicos
Fungicidas Benzimidazois
Foram introduzidos em 1970 e em 1977 a sarna apresentou resistência. Em SC foram encontrados isolados tolerantes a 1.000 ppm. A resistência a esses fungicidas foram atribuídas a mutações nas posições 198 e 200 da B-tubulina, assim o fungicida não conecta mais. Os isolados podem ser: altamente resistentes, muito resistentes e moderadamente resistentes. Ex: Corynespora cassiicola X Carbendazim.
Fungicidas DMIs (Inibidores da desmetilação)
Utilizado a partir da década de 1980. Inicialmente foram utiilzados triforine e bitertanol e após fenarimol, tebuconazol, etc. Quatro grupos para sarna (piperazinas, imidazois, pirimidinas e triazois) e mais de doze marcas. Mais de 20 anos de uso no Brasil, perda da eficiência no RS em SC. Necessidade de novo monitoramento.
Fungicidas QoI (Inibidores de quinona oxidase)
A introdução comercial ocorreu em 1996 e no mesmo ano foram detectados isolados resistentes de Podosphaera fusca (Fernández-Ortuño et al., 2006). A resistência é baseada em mutações no gene do citocromo b. A mutação no segmento G143A é o principal mecanismo de resistência, com 25 mutantes conhecidos. Essa mutação confere resistência completa, pois a presença de um intron impede duplicação DNA.
Ausência de controle pelo uso isolado de estrobilurinas:
Blumeria graminis, trigo e cevada
Mycosphaerella graminicola (= Septoria tritici), trigo Pyrenophora tritici-repentis, trigo
Plasmopara viticola, uva
Alternaria alternata, vários hospedeiros
Fungicidas QoI (Inibidores de quinona oxidase)
O segundo mecanismo de resistência é a mutação no segmento F129L e F137L, para a qual foram descritos 7 mutantes. Essa mutação confere resistência parcial. A Pyrenophora tritici-repentis [trigo]; Pyrenophora teres [cevada]; Alternaria solani [batata] apresentam controle reduzido pelo uso isolado de estrobilurinas. A mutação no segmento F137L é o terceiro mecanismo de resistência, descrito em Pyrenophora tritici-repentis, conferindo resistência parcial. Outros mecanismos de resistência identificados foram a respiração alternativa (Botrytis cinerea e Venturia inaequalis) e proteínas de efluxo situadas na membrana que bombeiam as estrubilurinas para fora da célula fúngica.
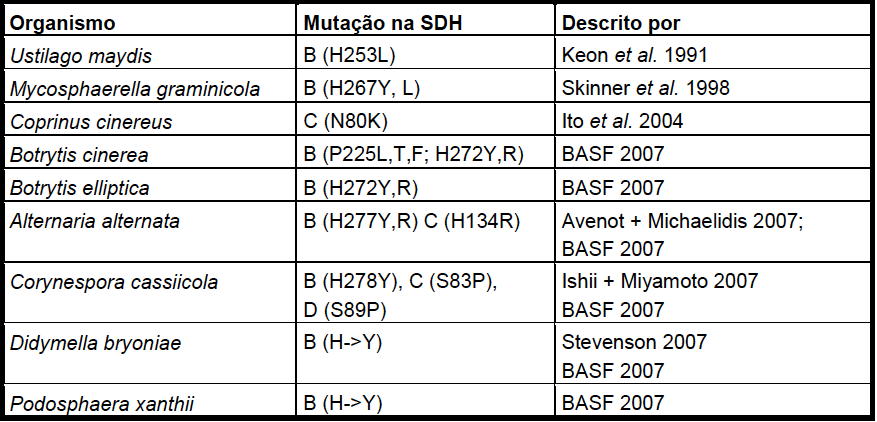
Fungicidas SDHI (Inibidores da Succinato Desidrogenase)
A resistência é qualitativa (abrupta) e o mecanismo ainda não é conhecido, supostamente uma alteração no gene que codifica a proteína alvo (SDHI).
Resistência a carboxamidas (SDHI):
REFERÊNCIAS BIBLIOGRÁFICAS
ARDUIM, G. D. S., REIS, E. M., BARCELLOS, A. L., & TURRA, C. (2012). In vivo sensitivity reduction of Puccinia triticina races, causal agent of wheat leaf rust, to DMI and QoI fungicides. Summa Phytopathologica, 38(4), 306-311.
VENOT, H. F., & MICHAILIDES, T. J. (2007). Resistance to boscalid fungicide in Alternaria alternata isolates from pistachio in California. Plant Disease, 91(10), 1345-1350.
BRENT, K. J., & HOLLOMON, D. W. (1998). Fungicide resistance: the assessment of risk (pp. 1-48). Brussels, Belgium: Global Crop Protection Federation.
DEISING, H. B.; REIMANN, S.; PASCHOLATI, S. F. Mechanisms and significance of fungicide resistance. Brazilian Journal of Microbiology, v. 39, n. 2, p. 286-295, 2008.
ELAD, Y., YUNIS, H., & KATAN, T. (1992). Multiple fungicide resistance to benzimidazoles, dicarboximides and diethofencarb in field isolates of Botrytis cinerea in Israel. Plant Pathology, 41(1), 41-46.
HARTMAN, G. L., MILES, M. R., & FREDERICK, R. D. (2005). Breeding for resistance to soybean rust. Plant disease, 89(6), 664-666.
KLOSOWSKI, A. C., MAY DE MIO, L. L., MIESSNER, S., RODRIGUES, R., & STAMMLER, G. (2015). Detection of the F129L mutation in the cytochrome b gene in Phakopsora pachyrhizi. Pest management science.
KOLLER, W., & WILCOX, W. F. (2001). Evidence for the predisposition of fungicide-resistant isolates of Venturia inaequalis to a preferential selection for resistance to other fungicides. Phytopathology, 91(8), 776-781.
MA, Z., & MICHAILIDES, T. J. (2005). Advances in understanding molecular mechanisms of fungicide resistance and molecular detection of resistant genotypes in phytopathogenic fungi. Crop Protection, 24(10), 853-863.
RUSSELL, P. E. (2002). Sensitivity baselines in fungicide resistance research and management (pp. 1-53). Brussels: Crop Life International.
SCHNEIDER, R. W., HOLLIER, C. A., WHITAM, H. K., PALM, M. E., MCKEMY, J. M., HERNANDEZ, J. R., … & DEVRIES-PATERSON, R. (2005). First report of soybean rust caused by Phakopsora pachyrhizi in the continental United States. Plant disease, 89(7), 774-774.
SENSEMAN, S. A. (Ed.). Herbicide handbook. 9.ed. Lawrence: Weed Science Society of America, 2007. 458 p.
STAUB, T. (1991). Fungicide resistance: practical experience with antiresistance strategies and the role of integrated use. Annual review of phytopathology, 29(1), 421-442.
VIDAL, R.A. Ação dos herbicidas. Porto Alegre: Edição do Autor, 2002. 89 p.




